
Trypanocidal and Anti-Inflammatory Effects of Three ent-Kaurane Diterpenoids from Gymnocoronis spilanthoides var. subcordata (Asteraceae)
 , , , , and
, , , , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Isolation and Purification of Phytochemicals from G. spilanthoides var. subcordata
2.3. Purity of the Isolated Compounds
2.4. Structure Elucidation of the Isolated Compounds
2.5. Trypanosoma cruzi In Vitro Testing
2.5.1. Parasites
2.5.2. In Vitro Activity Assay against Epimastigotes
2.5.3. In Vitro Activity Assay against Amastigotes
2.5.4. In Vitro Activity Assay against Trypomastigotes
2.6. Cytotoxicity Assay
2.7. Transmission Electron Microscopy
2.8. Anti-Inflammatory Properties of Compound 1
2.8.1. Effect of Compound 1 on the Type-I Interferon Pathway
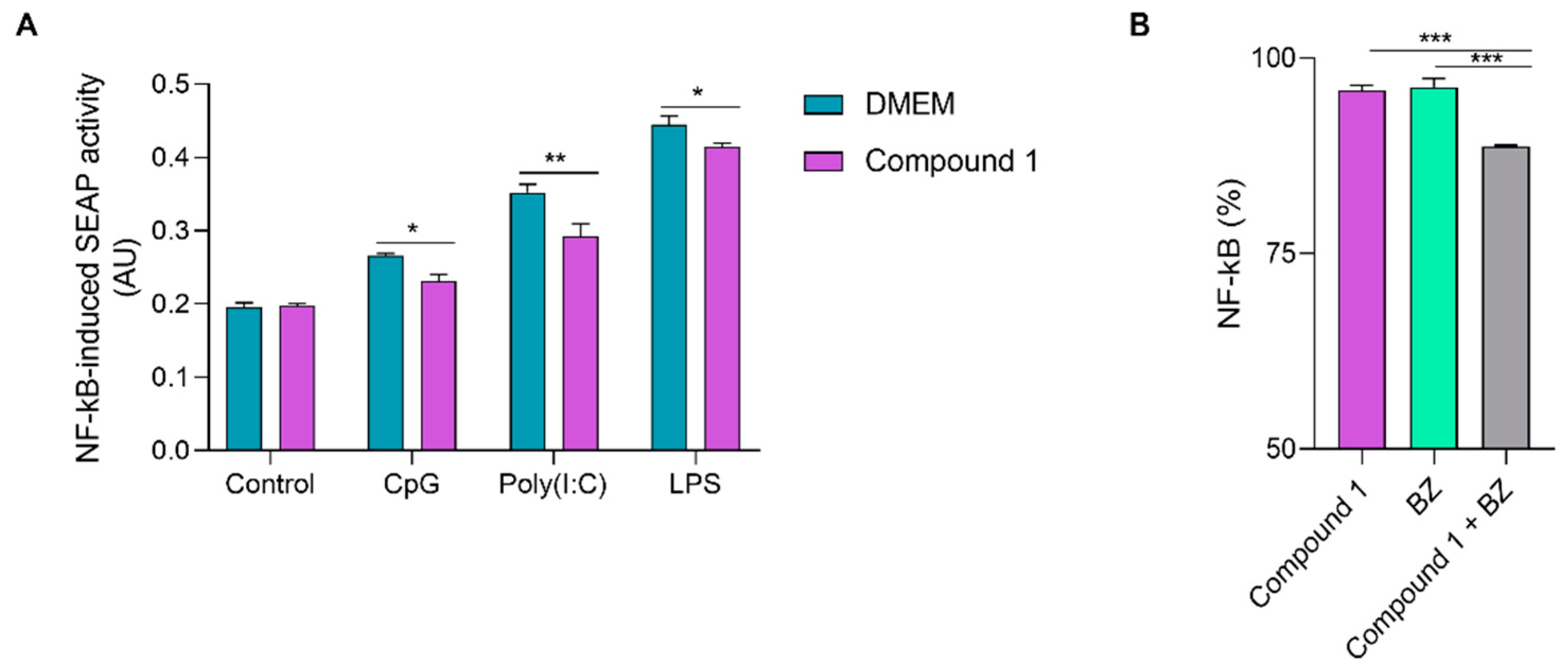
2.8.2. Effect of Compound 1 on the Secretion of NF-kB
2.8.3. Nitric Oxide Production in the Presence of Compound 1
2.9. Trypanosoma cruzi In Vivo Testing
2.9.1. Mice
2.9.2. In Vivo Trypanocidal Activity
2.10. Statistical Analysis
3. Results
3.1. Isolated Phytochemicals from G. spilanthoides var. subcordata
3.2. In Vitro Assays against Trypanosoma cruzi
3.3. Cytotoxicity and Selectivity against Trypanosoma cruzi
3.4. Transmission Electron Microscopy
3.5. Anti-Inflammatory Activity of Compound 1
3.6. In Vivo Assay in T. cruzi Murine Model
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- La Enfermedad de Chagas (Tripanosomiasis Americana). Available online: https://www.who.int/es/news-room/fact-sheets/detail/chagas-disease-(american-trypanosomiasis) (accessed on 12 July 2023).
- PAHO/WHO|Pan American Health Organization. Chagas Disease. Available online: https://www.paho.org/en/topics/chagas-disease (accessed on 22 November 2023).
- López-Vélez, R.; Norman, F.F.; Bern, C. 103—American Trypanosomiasis (Chagas Disease). In Hunter’s Tropical Medicine and Emerging Infectious Diseases, 10th ed.; Ryan, E.T., Hill, D.R., Solomon, T., Aronson, N.E., Endy, T.P., Eds.; Elsevier: London, UK, 2020; pp. 762–775. ISBN 978-0-323-55512-8. [Google Scholar]
- CDC-DPDx. American Trypanosomiasis. Available online: https://www.cdc.gov/dpdx/trypanosomiasisamerican/index.html (accessed on 11 February 2024).
- Morillo, C.A.; Marin-Neto, J.A.; Avezum, A.; Sosa-Estani, S.; Rassi, A.; Rosas, F.; Villena, E.; Quiroz, R.; Bonilla, R.; Britto, C.; et al. Randomized Trial of Benznidazole for Chronic Chagas’ Cardiomyopathy. N. Engl. J. Med. 2015, 373, 1295–1306. [Google Scholar] [CrossRef] [PubMed]
- Pecoul, B.; Batista, C.; Stobbaerts, E.; Ribeiro, I.; Vilasanjuan, R.; Gascon, J.; Pinazo, M.J.; Moriana, S.; Gold, S.; Pereiro, A.; et al. The BENEFIT Trial: Where Do We Go from Here? PLoS Negl. Trop. Dis. 2016, 10, e0004343. [Google Scholar] [CrossRef] [PubMed]
- DNDi América Latina. Enfermedad de Chagas. Available online: https://dndial.org/es/doencas/doenca-de-chagas/ (accessed on 22 November 2023).
- Gravina, H.D.; Antonelli, L.; Gazzinelli, R.T.; Ropert, C. Differential Use of TLR2 and TLR9 in the Regulation of Immune Responses during the Infection with Trypanosoma cruzi. PLoS ONE 2013, 8, e63100. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. Signaling to NF-κB by Toll-like Receptors. Trends Mol. Med. 2007, 13, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Cruz, J.S.; Machado, F.S.; Ropert, C.; Roman-Campos, D. Molecular Mechanisms of Cardiac Electromechanical Remodeling during Chagas Disease: Role of TNF and TGF-β. Trends Cardiovasc. Med. 2017, 27, 81–91. [Google Scholar] [CrossRef]
- Grijalva, A.; Gallo Vaulet, L.; Agüero, R.N.; Toledano, A.; Risso, M.G.; Quarroz Braghini, J.; Sosa, D.; Ruybal, P.; Repetto, S.; Alba Soto, C.D. Interleukin 10 Polymorphisms as Risk Factors for Progression to Chagas Disease Cardiomyopathy: A Case-Control Study and Meta-Analysis. Front. Immunol. 2022, 13, 946350. [Google Scholar] [CrossRef] [PubMed]
- Rolski, F.; Błyszczuk, P. Complexity of TNF-α Signaling in Heart Disease. J. Clin. Med. 2020, 9, 3267. [Google Scholar] [CrossRef]
- Nasim, N.; Sandeep, I.S.; Mohanty, S. Plant-Derived Natural Products for Drug Discovery: Current Approaches and Prospects. Nucleus 2022, 65, 399–411. [Google Scholar] [CrossRef]
- Barbosa, H.; Thevenard, F.; Quero Reimão, J.; Tempone, A.G.; Honorio, K.M.; Lago, J.H.G. The Potential of Secondary Metabolites from Plants as Drugs or Leads against Trypanosoma cruzi—An Update from 2012 to 2021. Curr. Top. Med. Chem. 2023, 23, 159–213. [Google Scholar] [CrossRef]
- Chatelain, E. Chagas Disease Drug Discovery: Toward a New Era. SLAS Discov. 2015, 20, 22–35. [Google Scholar] [CrossRef]
- Hurrell, J.; Bazzano, D.; Delucchi, G. Biota Rioplatense. Volumen XI. Dicotiledoneas Herbáceas 1. Nativas y Exóticas; Editorial L.O.L.A. (Literature of Latin América): Buenos Aires, Argentina, 2006; pp. 124–125, 150–151, 164. [Google Scholar]
- Boppré, M.; Colegate, S.M. Recognition of Pyrrolizidine Alkaloid Esters in the Invasive Aquatic Plant Gymnocoronis spilanthoides (Asteraceae). Phytochem. Anal. 2015, 26, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Ardenghi, N.M.G.; Barcheri, G.; Ballerini, C.; Cauzzi, P.; Guzzon, F. Gymnocoronis spilanthoides (Asteraceae, Eupatorieae), a New Naturalized and Potentially Invasive Aquatic Alien in S Europe. Willdenowia 2016, 46, 265–273. [Google Scholar] [CrossRef]
- Selener, M.G.; Elso, O.; Grosso, C.; Borgo, J.; Clavin, M.; Malchiodi, E.L.; Cazorla, S.I.; Flavia, F.; Sülsen, V.P. Anti-Trypanosoma cruzi Activity of Extracts from Argentinean Asteraceae Species. Iran. J. Pharm. Res. 2019, 18, 1854–1861. [Google Scholar] [CrossRef] [PubMed]
- Elso, O.G.; Clavin, M.; Hernandez, N.; Sgarlata, T.; Bach, H.; Catalan, C.A.N.; Aguilera, E.; Alvarez, G.; Sülsen, V.P. Antiprotozoal Compounds from Urolepis hecatantha (Asteraceae). Evid. Based Complement. Altern. Med. 2021, 2021, e6622894. [Google Scholar] [CrossRef] [PubMed]
- Borgo, J.; Elso, O.G.; Gomez, J.; Coll, M.; Catalán, C.A.N.; Mucci, J.; Alvarez, G.; Randall, L.M.; Barrera, P.; Malchiodi, E.L.; et al. Anti-Trypanosoma cruzi Properties of Sesquiterpene Lactones Isolated from Stevia spp.: In Vitro and In Silico Studies. Pharmaceutics 2023, 15, 647. [Google Scholar] [CrossRef]
- Sülsen, V.P.; Cazorla, S.I.; Frank, F.M.; Di Leo Lira, P.M.R.; Anesini, C.A.; GutierrezYapu, D.; GiménezTurba, A.; Bandoni, A.L.; Malchiodi, E.L.; Muschietti, L.V.; et al. In Vitro Antiprotozoal Activity and Chemical Composition of Ambrosia Tenuifolia and A. Scabra Essential Oils. Nat. Prod. Commun. 2008, 3, 557–562. [Google Scholar] [CrossRef]
- Sánchez Alberti, A.; Beer, M.F.; Cerny, N.; Bivona, A.E.; Fabian, L.; Morales, C.; Moglioni, A.; Malchiodi, E.L.; Donadel, O.J.; Sülsen, V.P. In Vitro, In Vivo, and In Silico Studies of Cumanin Diacetate as a Potential Drug against Trypanosoma cruzi Infection. ACS Omega 2022, 7, 968–978. [Google Scholar] [CrossRef]
- Brengio, S.D.; Belmonte, S.A.; Guerreiro, E.; Giordano, O.S.; Pietrobon, E.O.; Sosa, M.A. The sesquiterpene lactone dehydroleucodine (DhL) affects the growth of cultured epimastigotes of Trypanosoma cruzi. J. Parasitol. 2000, 86, 407–412. [Google Scholar] [CrossRef]
- Sülsen, V.P.; Frank, F.M.; Cazorla, S.I.; Barrera, P.; Freixa, B.; Vila, R.; Sosa, M.A.; Malchiodi, E.L.; Muschietti, L.V.; Martino, V.S. Psilostachyin C: A Natural Compound with Trypanocidal Activity. Int. J. Antimicrob. Agents 2011, 37, 536–543. [Google Scholar] [CrossRef]
- RAW-LuciaTM ISG Cells. Available online: https://www.invivogen.com/raw-lucia-isg (accessed on 21 November 2023).
- Sun, J.; Zhang, X.; Broderick, M.; Fein, H. Measurement of Nitric Oxide Production in Biological Systems by Using Griess Reaction Assay. Sensors 2003, 3, 276–284. [Google Scholar] [CrossRef]
- Institute of Laboratory Animal Resources (US); Committee on Care and Use of Laboratory Animals. Guide for the Care and Use of Laboratory Animals: Eighth Edition; National Academies Press: Washington, DC, USA, 2010; ISBN 978-0-309-18663-6. [Google Scholar]
- Shimizu, S.; Miyase, T.; Umehara, K.; Ueno, A. Kaurane-Type Diterpenes from Adenostemma Lavenia O. Kuntze. Chem. Pharm. Bull. 1990, 38, 1308–1312. [Google Scholar] [CrossRef]
- Herz, W.; Sharma, R.P. New Hydroxylated Ent-Kauranoic Acids from Eupatorium Album. J. Org. Chem. 1976, 41, 1021–1026. [Google Scholar] [CrossRef]
- Manarin, R.; Pascutti, M.F.; Ruffino, J.P.; De Las Heras, B.; Boscá, L.; Bottasso, O.; Revelli, S.; Serra, E. Benznidazole Blocks NF-κB Activation but Not AP-1 through Inhibition of IKK. Mol. Immunol. 2010, 47, 2485–2491. [Google Scholar] [CrossRef]
- Varela, J.; Serna, E.; Torres, S.; Yaluff, G.; De Bilbao, N.; Miño, P.; Chiriboga, X.; Cerecetto, H.; González, M. In Vivo Anti-Trypanosoma cruzi Activity of Hydro-Ethanolic Extract and Isolated Active Principles from Aristeguietia glutinosa and Mechanism of Action Studies. Molecules 2014, 19, 8488–8502. [Google Scholar] [CrossRef] [PubMed]
- Bohlmann, F.; Mahanta, P.K. Kaurenic Acid Derivatives from Adenostemma caffrum. Phytochemistry 1978, 17, 814–815. [Google Scholar] [CrossRef]
- Cheng, P.C.; Hufford, C.D.; Doorenbos, N.J. Isolation of 11-Hydroxyated Kauranic Acids from Adenostemma lavenia. J. Nat. Prod. 1979, 42, 183–186. [Google Scholar] [CrossRef] [PubMed]
- Sosa, A.M.; Amaya, S.; Salamanca Capusiri, E.; Gilabert, M.; Bardón, A.; Giménez, A.; Vera, N.R.; Borkosky, S.A. Active Sesquiterpene Lactones against Leishmania amazonensis and Leishmania braziliensis. Nat. Prod. Res. 2016, 30, 2611–2615. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Guo, Y.; Gao, Y.; Wang, S.; Wang, X.; Xie, Z.; Niu, H.; Chang, W.; Liu, L.; Yuan, H.; et al. Ent-Kaurane Diterpenoids from Chinese Liverworts and Their Antitumor Activities through Michael Addition as Detected In Situ by a Fluorescence Probe. J. Med. Chem. 2015, 58, 3944–3956. [Google Scholar] [CrossRef]
- Barros de Alencar, M.V.O.; de Castro e Sousa, J.M.; Rolim, H.M.L.; de Medeiros, M.d.G.F.; Cerqueira, G.S.; de Castro Almeida, F.R.; Citó, A.M.d.G.L.; Ferreira, P.M.P.; Lopes, J.A.D.; de Carvalho Melo-Cavalcante, A.A.; et al. Diterpenes as Lead Molecules against Neglected Tropical Diseases. Phytother. Res. 2017, 31, 175–201. [Google Scholar] [CrossRef]
- Izumi, E.; Ueda-Nakamura, T.; Veiga, V.F., Jr.; Pinto, A.C.; Nakamura, C.V. Terpenes from Copaifera Demonstrated In Vitro Antiparasitic and Synergic Activity. J. Med. Chem. 2012, 55, 2994–3001. [Google Scholar] [CrossRef]
- Menna-Barreto, R.F.S. Cell Death Pathways in Pathogenic Trypanosomatids: Lessons of (over)Kill. Cell Death Dis. 2019, 10, 93. [Google Scholar] [CrossRef]
- De Souza, W. From the Cell Biology to the Development of New Chemotherapeutic Approaches against Trypanosomatids: Dreams and Reality. Kinetoplastid Biol. Dis. 2002, 1, 3. [Google Scholar] [CrossRef]
- Cristovão-Silva, A.C.; Brelaz-de-Castro, M.C.A.; Hernandes, M.Z.; Pereira, V.R.A. Chagas Disease: Immunology of the Disease at a Glance. Cytokine Growth Factor Rev. 2021, 62, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Vellozo, N.S.; Matos-Silva, T.C.; Lopes, M.F. Immunopathogenesis in Trypanosoma cruzi Infection: A Role for Suppressed Macrophages and Apoptotic Cells. Front. Immunol. 2023, 14, 1244071. [Google Scholar] [CrossRef] [PubMed]
- Chessler, A.-D.C.; Caradonna, K.L.; Da’dara, A.; Burleigh, B.A. Type I Interferons Increase Host Susceptibility to Trypanosoma cruzi Infection. Infect. Immun. 2011, 79, 2112–2119. [Google Scholar] [CrossRef]
- Gil-Jaramillo, N.; Rocha, A.P.; Raiol, T.; Motta, F.N.; Favali, C.; Brigido, M.M.; Bastos, I.M.D.; Santana, J.M. The First Contact of Human Dendritic Cells with Trypanosoma cruzi Reveals Response to Virus as an Unexplored Central Pathway. Front. Immunol. 2021, 12, 638020. [Google Scholar] [CrossRef] [PubMed]
- Nurlela, N.; Badrunanto; Ilmiawati, A.; Nurcholis, W.; Takemori, H.; Batubara, I. The Medicinal Potential of Plants from the Adenostemma genus. J. Appl. Pharm. Sci. 2023, 13, 1–11. [Google Scholar] [CrossRef]
- Waghabi, M.C.; Coutinho-Silva, R.; Feige, J.-J.; Higuchi, M.d.L.; Becker, D.; Burnstock, G.; de Araújo-Jorge, T.C. Gap Junction Reduction in Cardiomyocytes Following Transforming Growth Factor-β Treatment and Trypanosoma cruzi Infection. Mem. Inst. Oswaldo Cruz 2009, 104, 1083–1090. [Google Scholar] [CrossRef]
- Santos, E.d.S.; Silva, D.K.C.; dos Reis, B.P.Z.C.; Barreto, B.C.; Cardoso, C.M.A.; Ribeiro dos Santos, R.; Meira, C.S.; Soares, M.B.P. Immunomodulation for the Treatment of Chronic Chagas Disease Cardiomyopathy: A New Approach to an Old Enemy. Front. Cell. Infect. Microbiol. 2021, 11, 765879. [Google Scholar] [CrossRef]
- Rodrigues, M.M.; Oliveira, A.C.; Bellio, M. The Immune Response to Trypanosoma cruzi: Role of Toll-Like Receptors and Perspectives for Vaccine Development. J. Parasitol. Res. 2012, 2012, e507874. [Google Scholar] [CrossRef]
- Mendes da Silva, L.D.; Gatto, M.; Miziara de Abreu Teodoro, M.; de Assis Golim, M.; Pelisson Nunes da Costa, É.A.; Capel Tavares Carvalho, F.; Ramos Rodrigues, D.; Câmara Marques Pereira, P.; Victoriano de Campos Soares, Â.M.; Calvi, S.A. Participation of TLR2 and TLR4 in Cytokines Production by Patients with Symptomatic and Asymptomatic Chronic Chagas Disease. Scand. J. Immunol. 2017, 85, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Villanueva-Lizama, L.E.; Cruz-Chan, J.V.; Versteeg, L.; Teh-Poot, C.F.; Hoffman, K.; Kendricks, A.; Keegan, B.; Pollet, J.; Gusovsky, F.; Hotez, P.J.; et al. TLR4 Agonist Protects against Trypanosoma cruzi Acute Lethal Infection by Decreasing Cardiac Parasite Burdens. Parasite Immunol. 2020, 42, e12769. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, L.M.D.; Gollob, K.J.; Zingales, B.; Dutra, W.O. Pathogen Diversity, Immunity, and the Fate of Infections: Lessons Learned from Trypanosoma cruzi Human–Host Interactions. Lancet Microbe 2022, 3, e711–e722. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | δ 1H (J Hz) * | δ 13C (HSQC) | H-H COSY | NOESY | HMBC | Reported Ʊ Pyridine-d5 |
|---|---|---|---|---|---|---|
| 1a a 1b | 1.86ddd (12, 12, 4) 1.22 d br (12) | 31.6 CH2 | 1b, 2a, 2b 1a, 2a, 2b, 3a (W) | 1b 1a, 11 | 20, 3a | 32.7 ¥ |
| 2a 2b | 1.79 m 1.34 m | 18.5 CH2 | 1a,1b, 2b, 3a, 3b 1a, 1b, 2a, 3a, 3b | --- --- | 1a, 3b | 19.8 |
| 3a 3b | 1.96 d br (13) 0.93 td (13, 4) | 37.4 CH2 | 2a, 2b, 3b, 1b (W) 2a, 2b, 3a | 3b, 18 3a | 18, 1b | 38.6 |
| 4 | --- | 42.9 C | --- | --- | 18, 3b | 44.5 |
| 5 | 1.77 (overlapped m) | 48.6 CH | 6a,b | --- | 20, 18, 3a, 7a, 7b | 50.0 |
| 6a 6b | 1.72–1.82 (overlapped m) | 20.0 CH2 | 5, 6b, 7a, 7b 5, 6a, 7a, 7b | --- --- | 7a, 5 | 21.2 |
| 7a | 2.13 m | 29.6 CH2 | 6a,b, 7b | --- | 5, 6a,b | 31.0 ¥ |
| 7b | 0.97 m | 6a,b, 7a | --- | |||
| 8 | --- | 55.7 C | --- | --- | 14a, 7a, 7b, 14b, 6a,b | 56.9 |
| 9 | --- | 77.9 C | --- | --- | 20, 14a, 14b, 12b, 11 | 79.5 |
| 10 | --- | 43.4 C | --- | --- | 20, 6a,b | 44.4 |
| 11 | 3.77 dd br (3.8, 2) | 64.8 CH | 12a,C11-OH | 1b, 12a, 12b | 13, 12b | 66.3 |
| 12a | 2.13 m | 41.6 CH2 | 11, 13, 12b | --- | 14a, 14b | 42.7 |
| 12b | 2.06 d br (14) | 11, 13, 12a, 14b (W) | 12a, 11, 13 | |||
| 13 | 2.92 m | 35.9 CH | 12a, 12b, 14b | 14a,12a, 12b | 14a, 17a, 17b | 37.4 |
| 14a | 2.53 d (12) | 37.3 CH2 | 14b | 20, 14b | 12a, 7a | 38.6 ¥ |
| 14b | 1.32ddd (12, 4, 3) | 13, 14a, 12b (W) | 14a | |||
| 15 | --- | 206.1 C | --- | --- | 14a, 17a, 17b | 207.3 |
| 16 | --- | 150.4 C | --- | --- | 12a, 14a, 17a, 17b | 151.6 |
| 17a | 5.52 s | 110.8 CH2 | 17b | 17b | 14a | 111.1 |
| 17b | 5.15 s | 17a | 17a | |||
| 18 b | 1.12 s | 28.8 CH3 | --- | --- | 29.7 | |
| 19 | --- | 178.9 C | --- | --- | 3a, 5, 18 | 180.4 |
| 20 b | 1.01 s | 16.6 CH3 | --- | 2a, 14a | 1a, 5 | 17.6 |
| Position | Compound 2 | Compound 3 | ||
|---|---|---|---|---|
| δ C | δ H (J in Hz) | δ C | δ H (J in Hz) | |
| 1 | 39.7 t | 39.5 t | ||
| 2 | 18.8 t | 18.7 t | ||
| 3 | 37.7 t | 37.7 t | ||
| 4 | 43.6 s | 43.6 s | ||
| 5 | 55.9 d | 1.25 m * | 56.0 d | |
| 6 | 19.9 t | 20.0 t | ||
| 7 | 36.6 t | 37.2 t | ||
| 8 | 50.5 s | 51.0 s | ||
| 9 | 63.1 d | 1.40 s br | 62.9 d | |
| 10 | 39.0 s | 38.7 s | ||
| 11 | 66.3 d | 4.05 d br (4.8) | 65.3 d | 3.93 d br (5.6) |
| 12a | 41.2 t | 2.11 ddd (14.5, 4.8, 3.1) | 33.2 t | |
| 12b | 1.98 m * | |||
| 13 | 36.9 d | 3.06 m | 34.8 d | 2.44 m |
| 14a | 33.7 t | 2.37 d (12) | 34.5 t | 2.35 d (12) |
| 14b | 1.42 * | |||
| 15 | 209.6 s | 222.5 s | ||
| 16 | 150.2 s | 49.5 d | 2.28 quint (6.8) | |
| 17a | 113.0 t | 5.87 s | 11.1 q | 1.25 § d (6.8) |
| 17b | 5.27 s | |||
| 18 § | 28.9 q | 1.28 s | 28.8 q | 1.26 s |
| 19 | 181.1 s | 176.3 s | ||
| 20 § | 15.6 q | 0.95 s | 15.3 q | 0.92 s |
| Compounds | IC50 ± SEM µg/mL (µM) | |
|---|---|---|
| Epimastigotes | Amastigotes | |
| 1 | 3.7 ± 0.5 (10.6) | 2.1 ± 0.3 (6.1) |
| 2 | 5.2 ± 0.3 (15.9) | 6.5 ± 0.2 (19.5) |
| 3 | 1.6 ± 0.6 (4.8) | 20.1 ± 8.2 (60.6) |
| Benznidazole | 1.08 ± 0.2 (4.2) | 0.5 ± 0.04 (1.9) |
| Compound | CC50 | SI | ||
|---|---|---|---|---|
| µg/mL ± SEM | µM | Epimastigotes | Amastigotes | |
| 1 | 112.0 ± 3.4 | 321.8 | 30.3 | 52.7 |
| 2 | 7.8 ± 0.2 | 23.3 | 1.5 | 1.2 |
| 3 | 4.9 ± 1.6 | 14.8 | 3.1 | 0.2 |
| Benznidazole | 45.3 ± 1.8 | 81.5 | 19.6 | 42.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Selener, M.G.; Borgo, J.; Sarratea, M.B.; Delfino, M.A.; Laurella, L.C.; Cerny, N.; Gomez, J.; Coll, M.; Malchiodi, E.L.; Bivona, A.E.; et al. Trypanocidal and Anti-Inflammatory Effects of Three ent-Kaurane Diterpenoids from Gymnocoronis spilanthoides var. subcordata (Asteraceae). Pharmaceutics 2024, 16, 415. https://doi.org/10.3390/pharmaceutics16030415
Selener MG, Borgo J, Sarratea MB, Delfino MA, Laurella LC, Cerny N, Gomez J, Coll M, Malchiodi EL, Bivona AE, et al. Trypanocidal and Anti-Inflammatory Effects of Three ent-Kaurane Diterpenoids from Gymnocoronis spilanthoides var. subcordata (Asteraceae). Pharmaceutics. 2024; 16(3):415. https://doi.org/10.3390/pharmaceutics16030415
Chicago/Turabian StyleSelener, Mariana G., Jimena Borgo, Maria Belen Sarratea, Maria Alicia Delfino, Laura C. Laurella, Natacha Cerny, Jessica Gomez, Mauro Coll, Emilio L. Malchiodi, Augusto E. Bivona, and et al. 2024. "Trypanocidal and Anti-Inflammatory Effects of Three ent-Kaurane Diterpenoids from Gymnocoronis spilanthoides var. subcordata (Asteraceae)" Pharmaceutics 16, no. 3: 415. https://doi.org/10.3390/pharmaceutics16030415